首页 / 产品和服务 / 产品类型 / 抗体产品 / 特色抗体推荐
Wnt/β-catenin signaling is an evolutionarily conserved pathway which is involved in embryonic development and adult homeostasis, first reported in 1982 with identification of mouse proto-oncogene int1 (also known as Wnt1). Later the homolog of int1, Wingless (Wg) from Drosophila melanogaster, was identified to promote wing development in fruit flies[1, 2]. Since then, accumulative studies have reported crucial roles of the Wnt/β-catenin signaling in development of various organs and tissues(heart, liver, lungs, limbs and so on)[3, 4] and adult homeostasis such as regeneration of skin, gut, hair and bone marrow [5-8]. On the other hand, abnormal Wnt/β-catenin signaling can lead to developmental malformations and is associated with many types of disease including fibrosis, gastric and colorectal tumors, melanomas and hepatocellular carcinomas[9-15].
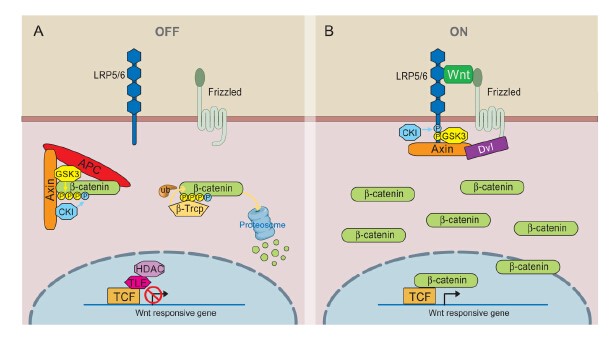
(1)Wnt-off state
β-catenin phosphorylation/degradation by the Axin complex
The phosphorylation or degradation of cytosolic β-catenin by Wnt are the essence of Wnt signaling. The scaffolding protein Axin interacts with GSK3, APC and β-catenin with sequential phosphorylation of β-catenin[16]. The E3 ubiquitin ligase β-Trcp binds to phosphorylated β-catenin leading to β-catenin ubiquitination and degradation. Several aspects of the Axin complex deserve further discussion. These core Axin complex components[16] share a common goal of ensuring β-catenin phosphorylation and degradation. Besides, two abundant serine/threonine phosphatases, PP1 and PP2A, which counteract the action of GSK3 and/or CK1 in the Axin complex lead to reduce β-catenin degradation[17, 18].
APC function and APC-Axin cross regulation
The biochemical nature of APC has been enigmatic. A recent study suggested that the overexpression of Axin causes β-catenin degradation even in cells lacking APC function[19]. Another paradoxical observation is that APC has a positive function in physiological and ectopic Wg/Wnt signaling through the promotion of Axin degradation[20, 21]. Conversely Axin can also facilitate APC degradation upon overexpression[22], constituting perhaps the other side of the Axin-APC regulation circuit. However, mechanisms for Axin and APC degradation have not been characterized.
(2)Wnt-on state
Activation of Wnt receptors
Wnt signaling requires both Fz and LRP6 (or LRP5), likely through a Wnt-induced Fz-LRP6 complex. Wnt-induced LRP6 phosphorylation is a key event in receptor activation[23], which is also important for Fz function[24]. In addition, Fz function is usually linked to Dsh/Dvl[25], a cytoplasmic scaffolding protein that may directly interact with Fz [26]. One model shows that Fz-Dvl recruitment of the Axin-GSK3 complex initiates LRP6 phosphorylation by GSK3[24].
Inhibition of β-catenin phosphorylation
How receptor activation leads to inhibition of β-catenin phosphorylation remains uncertain, and available data suggest possible parallel mechanisms. In the LRP6-centric view, as constitutively activated forms of LRP6 fully activate β-catenin signaling in an apparently Fz and Dvl-independent manner[27]. On the other hand, Dsh overexpression in Drosophila or recombinant Dvl in Xenopus egg extracts can activate β-catenin signaling presumably in the absence of Arrow/LRP6[28, 29], and so does a GPCR-Fz chimeric protein in response to the GPCR ligand[30]. These results suggest that two parallel branches need to operate together to counteract efficient β-catenin phosphorylation/degradation, whereas overactivation of either branch is sufficient to stabilize β-catenin.
Several biochemical mechanisms by which Wnt inhibits β-catenin phosphorylation have been suggested[16]. (i) Wnt-induced and Dvl-(and Gα) -dependent Axin-GSK3 (or β-catenin) dissociation[3, 31]; (ii) Inhibition of GSK3[32-35]; (iii) Axin degradation[36-38].
| Main factors | Wnt pathway |
|---|---|
| The upstream regulators | Wnt, LRP5/6, Frizzled |
| The core factors | Axin, APC, GSK-3β, CKI |
| The downstream effectors | β-catenin,TCF |
Wnt signaling is involved in numerous fundamental processes essential for embryonic development and normal adult homeostasis. The Wnt family is a highly evolutionarily conserved family of proteins as shown by ectopic expression of Wnt1 from Drosophila in organisms, causing serious developmental issues[39, 40]. The human Wnt family is composed of nineteen different cysteine-rich glycoproteins acting as ligands for more than 15 receptors or co-receptors[41], which is involved in many cellular functions essential for normal organ development including cell proliferation, survival, self-renewal/differentiation etc[42, 43].
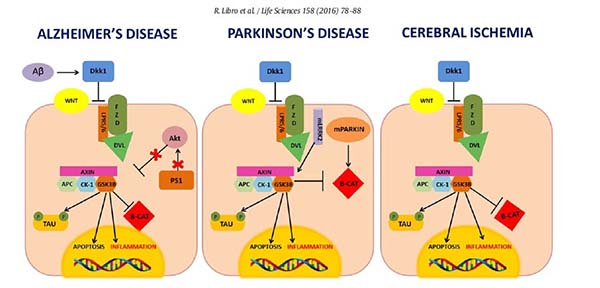
(1)Alzheimer’s disease
Alzheimer's disease (AD) is a progressive neurodegenerative disorder, characterized by the progressive loss of cholinergic neurons, resulting in mental decline and loss of cognitive processes including memory, judgment and reasoning. The main pathological hallmarks of AD are senile plaques, due to the insoluble Amyloid Beta (Aβ) aggregation and the Neurofibrillary Tangles (NFTs), formed by the hyperphosporylated microtubule-associated protein Tau[44].TheWnt signaling plays a central role in synaptic stability [45]as well as in the maintenance of blood-brain barrier (BBB) integrity [46].
(2)Cerebral ischemic injury
The cerebral ischemic injury can be caused by focal or global circulatory arrest in the brain[47]. The reduced cerebral blood flow in turn can cause oxidative stress and inflammation, which culminate in neuronal death [48]. In the peripheral blood of patients affected by cerebrovascular diseases increased levels of the Wnt agonist DKK1 were found and correlated with the inactivation of the Wnt canonical pathway[49]. Increased DKK1 and decreased β-catenin expression were observed also in a rat model of transient Focal Cerebral Ischemia (FCI) by Mastroiacovo et al. [50].
(3)Parkinson’s disease
Parkinson's disease (PD) is a neurodegenerative disorder characterized by the progressive loss of ventral midbrain dopaminergic (DA) neurons of the substantia nigra pars compacta causing typical motor symptoms, such as tremor and bradykinesia. Histologically, PD is characterized by intracellular inclusions enriched in α-synuclein aggregates, known as Lewy bodies. Several studies have suggested a link between PD pathogenesis and the Wnt canonical pathway. LRRK2 mutations in Roc, COR and kinase domains, are able to reduce the LRRK2-LRP6 binding affinity and have been associated with reduced activation of the Wnt canonical pathway and increased neurodegeneration. On the other hand, mutant LRRK2 possesses an increased ability to recruit GSK-3β. In turn, GSK-3β by phosphorylating the Tau protein, exacerbated neurodegeneration.
(1)Infarct healing
Myocardial infarction(MI) is the consequence of a partially or completely occluded coronary artery due to a thrombus, stenosis (due to atherosclerosis) or a coronary spasm. As a result blood flow is impaired and the myocardium is deprived from oxygen and nutrients. Wnt signaling is activated following MI in several cell types throughout the myocardium, suggesting an extensive role for Wnt signaling in cardiac repair.
(2)Cardiac hypertrophy
Cardiac hypertrophy is the response of the heart to an increased workload and is characterized by an increase in cardiomyocyte size. Dvl can be regarded as a critical factor in Wnt signaling that is able to induce cardiac hypertrophy when expressed in higher levels and engages both canonical and noncanonical pathways as well as Akt-mediated signaling. Moreover, both GSK3β knock-out and overexpression impair cardiogenesis[51] and the degree of GSK3β activity plays indeed a role in cardiac hypertrophy. On the other hand, Wnt5a activates noncanonical signaling and induces hypertrophy in cardiomyocyte cultures[52].
(3)Atherosclerosis
Indications for involvement of Wnt signaling in atherosclerosis are given by the presence of Wnt5a and DKK1 in murine and human lesions [53]. In addition, serum levels of DKK1 have been identified as an independent risk factor for the presence of calcification and atherosclerotic plaques in coronary arteries [54].
(1)Colorectal cancer
The mutational landscape of colorectal cancer shows genetic alterations in the Wnt pathway. Although the nature of these mutations may be distinct, they ultimately result in the stabilization of β-catenin, the key transducer of canonical Wnt signals. The most prevalent mutations are truncating events that inactivate the tumor suppressor Adenomatousis Polyposis Coli (APC)[55], a protein that binds and stabilizes AXIN2 and Glycogen synthase kinase 3β(GSK3β) to form the β-catenin destruction complex that leads to cytosolic β-catenin for proteasomal degradation. Wnt activating mutations occur early during colon tumorigenesis whereas the progression of the disease is often accompanied by other genetic alterations, most commonly seen in KRAS, BRAF, TP53, and SMAD4[56]. Thus, different context select for distinct mutational spectra, also with respect to Wnt signaling, and are accompanied with distinct biological and clinical presentation.
(2)Other cancers
The role of Wnt signaling in carcinogenesis extends beyond colorectal cancer. Mutations of pathway components are also frequently detected in a subset of hepatocellular carcinoma. There, hyperactivation of Wnt signaling is mediated by loss of function and inactivation of the Wnt negative regulators AXIN1 and/or AXIN2[57]. In addition, hyperactivation of Wnt signaling in breast cancers is rarely due to mutations in the pathway but rather a consequence of increased ligand production[58]. Another example of a Wnt-driven tumor is medulloblastoma, one of the most common juvenile brain tumors. At least 10% of all medulloblastoma are classified as the so-called Wnt subtype since they exhibit a high frequency of β-catenin activating mutations[59].
(1)Biological agents
Several biological agents have been studied in cancer by specifically targeting either aberrantly overexpressed Wnt receptors or the Wnt ligands. For instance, inhibition of Wnt/β-catenin signaling by adenovirus-mediated expression of Dickkopf-1 (Dkk-1), a potent secreted Wnt antagonist that interacts with the LRP5/6 co-receptors, has been shown to suppress epithelium proliferation in small intestine and colon[60, 61]. In addition, it has also been shown that Mesd, another LRP5/6 co-receptor inhibitor, effectively inhibits prostate cancer PC-3 cell proliferation in vitro and markedly decreases growth of breast cancer in the mouse mammary tumor virus-Wnt-1 transgenic mice[62, 63]. Moreover, large antibodies and small peptides have been developed to target the Wnt/β-catenin signaling as well. For instance, previous studies have demonstrated that monoclonal antibodies-neutralizing Wnt3a can suppress prostate tumor growth in a mouse model[64], and antibodies targeting Frizzled receptors are effective in various preclinical models including those of breast, colon and liver cancer[65, 66].
(2)Small molecule inhibitors of the Wnt/β-catenin pathway
Compared to biological agents, small molecule drugs exhibit several advantages including lower cost with greater ease of manufacturing, oral bioavailability and ability to penetrate into cells for intracellular targets. Here we classify the small molecule inhibitors of the Wnt/β-catenin signaling into three groups: small molecules that target cytoplasmic proteins, small molecules that target transcriptional factors and small molecules that target the co-activators.
(3)Small molecules that target cytoplasmic proteins of the Wnt/β-catenin cascade
Dvl: Three Dishevelled isoforms, Dvl1, Dvl2 and Dvl3, have been identified in mammalian species[67]. The special role of the Dvl PDZ domain in the Wnt pathway makes it an ideal pharmaceutical target. Three compounds (NSC 668036[68], J01-07a[69] and 3289-8625[70]) block the Wnt/β-catenin signaling by interacting with the groove of Dvl PDZ domain where the inhibitory Dapper proteins bind.
Tankyrase inhibitors: Tankyrase is a subgroup of Poly (ADP-ribose) polymerases (PARPs) family. Two isoforms of Tankyrase, Tankyrase 1(PARP5a) and Tankyrase 2 (PARP5b), have been identified to be involved in the Wnt/β-catenin signaling[71]. Both Tankyrase isoforms interact with and poly(ADPribosyl)ate Axin to stimulate Axin degradation through the ubiquitin-proteasome pathway[72]. A few small molecule Tankyrase inhibitors (IWR-1[73], XAV939[72], WIKI4, JW55, JW74 and G007-LK [74]) have been developed to inhibit the Wnt/β-catenin signaling.
Porcupine inhibitors: Porcupine is a membranebound O-acyltransferase (MBOAT) specific to Wnt post-translational acylation, which is required for subsequent Wnt secretion[75]. Targeting Porcupine for Wnt inhibition may represent a new therapeutic strategy for cancer therapy. There are some Porcupine inhibitor, such as IWP-2[76] and LGK974[77].
CK1 inhibitors: In the destruction complex, β-catenin is initially phosphorylated by CKIα at Ser45. This CKIα-mediated phosphorylation is a critical step for ubiquitin-dependent degradation of β-catenin. By screening libraries of FDA-approved drugs, Pyrvinium was identified with the ability to activate CK1α, leading to enhanced degradation of β-catenin[78]. Another CK1 kinase isoform, CK1ε, is involved in phosphorylation of E-cadherin upon Wnt binding to Frizzled-LRP5/6 receptor, leading to the release of β-catenin from its complex with E-cadherin and subsequent increase of the cellular threshold of free β-catenin. Recently Cheong and colleagues have identified a small molecule, PF670462, which can potently suppress the Wnt/β-catenin signaling by inhibiting CK1ε activity [79].
(1)Biological therapeutic agents
A number of biologic therapeutic agents targeting the Wnt pathway have entered clinical trials. The clinic Phase I trial of one of these agents, OMP18R5 (also known as Vantictumab, a fully humanized monoclonal antibody that targets frizzled receptor), has recently been completed by OncoMed Pharmaceuticals[80].
(2)Small molecule therapeutic agents
Other than biological agents, clinical trials of small molecule agents have also been conducted in recent years, too. For instance, Novartis Pharmaceuticals initiated a Phase I trial of the small molecule Porcupine inhibitor LGK974 for multiple malignancies (melanoma, breast cancer and pancreatic adenocarcinoma) that are associated with aberrant Wnt signaling. It is expected that the Phase I trial to obtain a maximum tolerated dose of LGK974 will be completed in January 2017. In addition, JW Pharmaceutical recently initiated a Phase I clinical study of CWP232291, a small molecule prodrug targeting β-catenin for degradation, in acute myeloid leukemia patients. The clinical trial is expected to be completed at the end of 2015, and no results have been disclosed publicly yet.
The evolutionarily conserved Wnt signaling pathway plays an indispensable role in regulating a diverse range of cellular activities. With the development of our understanding of Wnt signaling regulation and complexity is an increasing awareness of its role in physiological aspect and pathology, especially cancers. Although the Wnt signaling pathway could be the target for the cancer therapy, precise modulation of the Wnt pathway will be necessary to balance anti-tumor efficacy with adverse events and will be a challenge for ongoing and future clinical trials.





_IF_01.jpg?t=1783668060 "alt=")
_IF_02.jpg?t=1783668060 "alt=")











