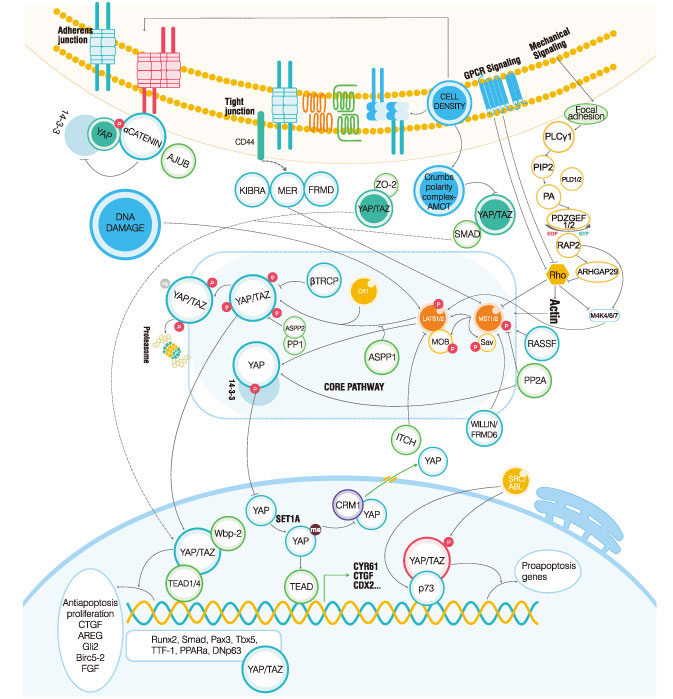
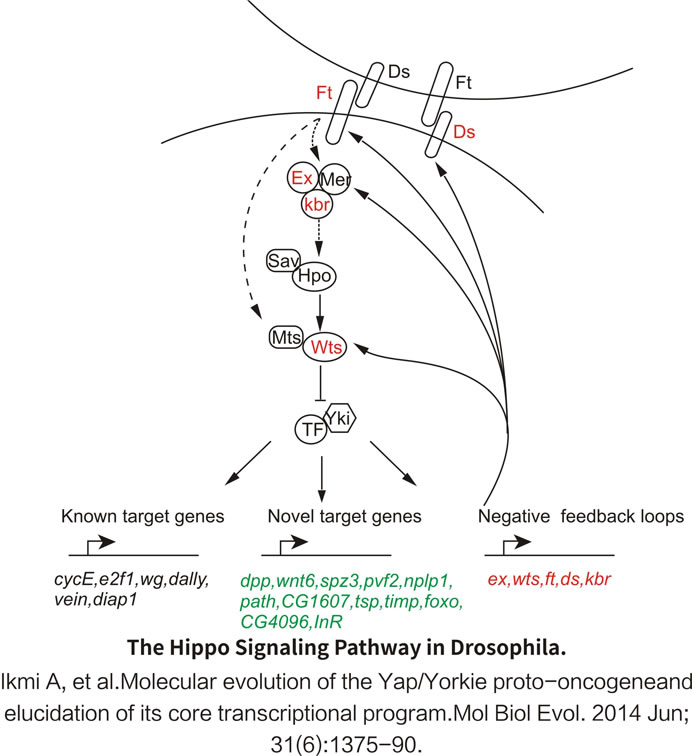
The Hippo pathway controls organ size by regulating cell proliferation, apoptosis, and stem cell self-renewal. Past research suggested that the dysregulation of the Hippo pathway may contribute to cancer development.
The Core of the Mammalian Hippo PathwayNot unlike the invertebrate cascade, in the mammalian Hippo pathway, the phosphorylation of the co-activators YAP and TAZ is caused by phosphorylation events via the Drosophila Mst ortholog, hippo and Wts (mammalian LATS1/2). Once phosphorylated, the protein kinase is activated, resulting in a series of changes in downstream processes.
Activation of the Mammalian Hippo PathwayYorkie (Yki) is a transcriptional co-activator; it cannot be directly combined with DNA, but needs to combine with the transcription factor Scalloped (Sd) to form the Yki-Sd complex, which plays a role in transcription regulation. The Yki-Sd transcription complex can be regulated by multiple genes related to cell proliferation and transcription regulation, such as E cyclin. When the organ reaches a certain size, the pathway is activated and phosphorylated (activated) Wts can be phosphorylated by Yki, and the 168th serine is activated. Yki binds to 14-3-3 after phosphorylation (inactivation). This process allows Yki to reside in the cytoplasm, which cannot form a transcriptional regulatory complex with Sd, and inhibit the transcription of downstream target genes.
Within the nucleus, both YAP and TAZ can activate gene transcriptions by binding to TEAD1/4. The activated genes (CTGF, AREG, BIRC5-2, FGF, and GLI-2) are required to promote cell growth and inhibit apoptosis. When phosphorylated by the SRC/ABL kinases, YAP and TAZ are then able to bind with p73 and activate proapoptotic gene transcriptions. Scientists have described more than one transcription factor as YAP/TAZ binding partners, but the functional significance of the factors require further research.

_WB_01.jpg?t=1779442213 "alt=")




_IP_02.jpg?t=1779442213 "alt=")